摘要
纤维素、半纤维素和木质素是木质纤维生物质细胞壁中的3种主要组分,其通过复杂的相互作用形成超分子网络结构,使木质纤维生物质具有优异的机械性能及抵抗物理、化学和微生物降解的能力,同时也导致生物质预处理和组分分离困难。本文总结了近年来关于纤维素、半纤维素与木质素间相互作用的研究进展,重点介绍了木质素与半纤维素之间的共价键、范德华力和静电作用等,以期在原子-分子水平上充分认识和了解细胞壁结构,为通过基因工程合成易降解的木质纤维原料,或寻找清洁高效的生物质精炼技术提供一定的理论指导。
木质纤维细胞壁是维持细胞一定形态、增加机械强度和防止外部侵害的一层硬质厚壁,具有由纤维素、半纤维素和木质素3种主要组分交联而成的复杂网络结构。其中,纤维素在植物细胞壁中含量最高,是由D-吡喃式葡萄糖通过β-1,4糖苷键连接而成的线性高分子多糖。数十年来,基原纤维(Elementary Microfibril)的主流假设模型是由纤维素分子链(18~36条)通过氢键作用组成,其直径为3~5 n
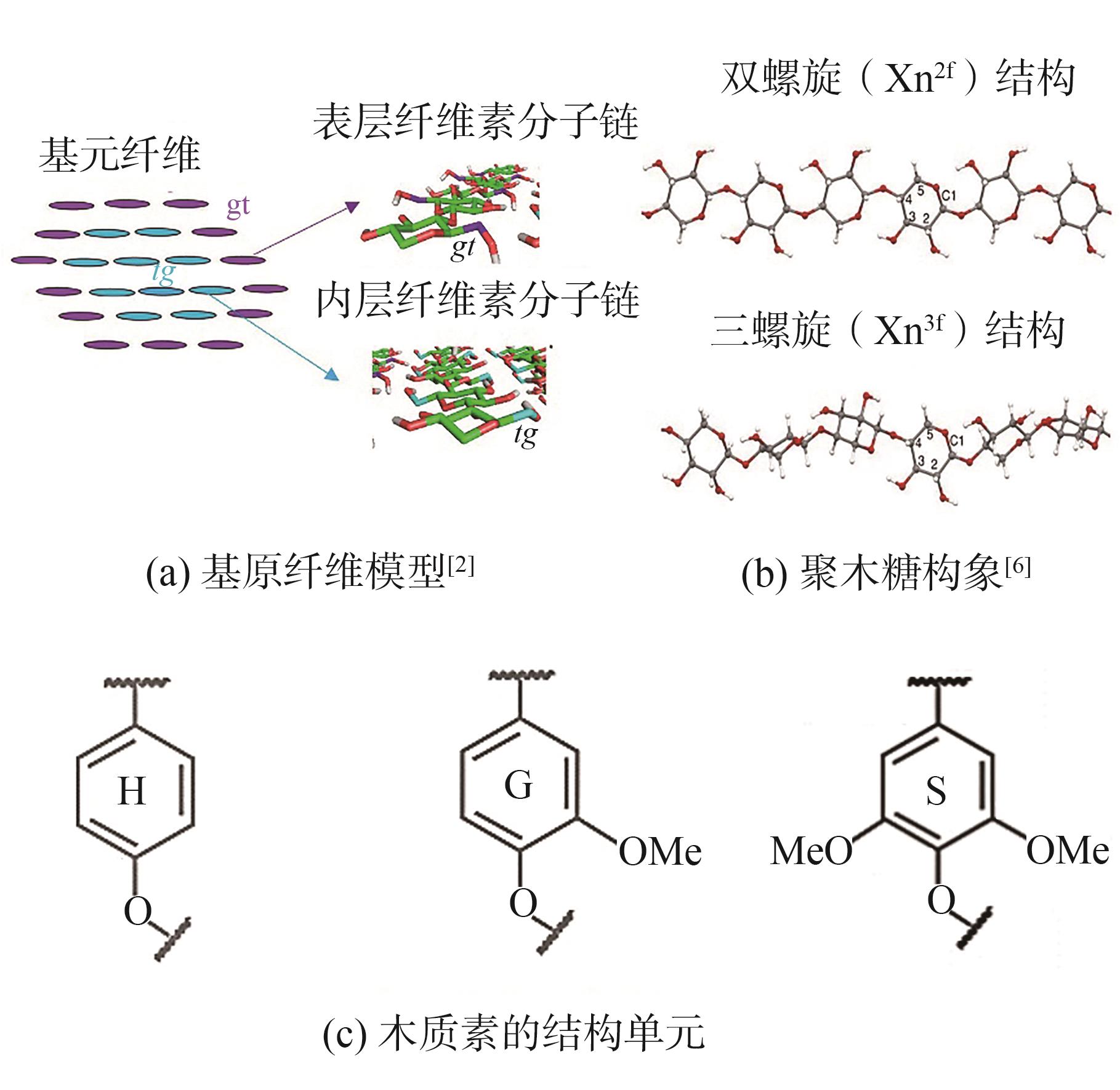
图1 植物细胞壁内主要组分的分子结构
Fig. 1 Molecular structure of main compositions in the cell wall of plants
木质纤维细胞壁中,纤维素、半纤维素和木质素3种主要组分间具有复杂的相互作用,这是其具有顽强的抗降解性的重要原因,也是当前生物质转化领域的研究热点与难点。由于主要组分的异质性及相互之间作用的复杂性,导致研究人员对生物质结构的原位表征和模型构建非常困难。目前,学者们主要通过透射电子显微镜(TEM)、核磁共振(NMR)和共聚焦显微拉曼光谱等技术,对细胞壁的分子信息进行解译,已取得了重要的进展。
本文综述了纤维素、半纤维素和木质素3种主要组分间的相互作用关系,特别是分子之间、亚纳米级上的原子信息和结构特点,以期更好地认识细胞壁超分子结构,为生物质组分的高效分离及转化提供一定的理论基础。
纤维素是细胞壁中含量最多的组分,约占细胞壁总质量的40%~50%。人们普遍认为,半纤维素填充在纤维素的周围,与纤维素主要通过氢键连接。近期,随着NMR技术的发展,学者们获得了更加详细的信息,发现拟南芥聚木糖主要以带状X
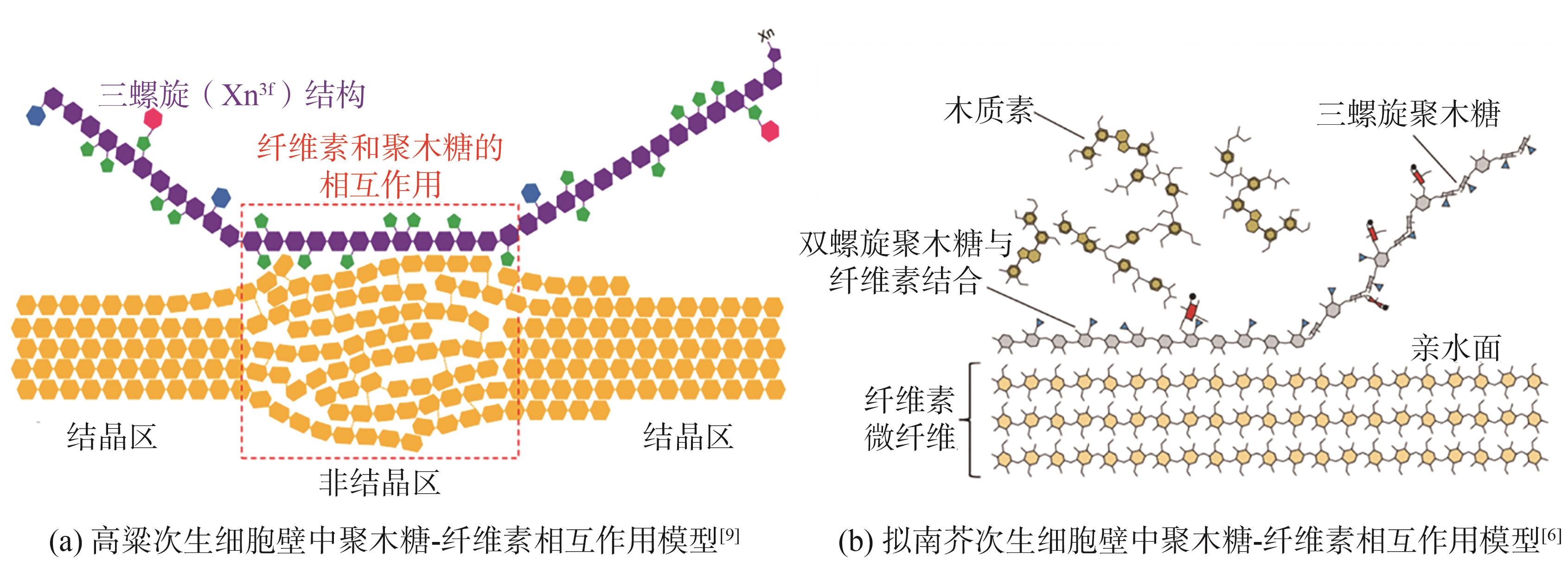
图2 聚木糖与纤维素之间相互作用关系模型
Fig. 2 Models of xylan-cellulose interaction
纤维素与木质素间也存在相互作用。尽管木质素是无定形的,但其与细胞壁的多糖物质纤维素及半纤维素以可控的方式连接,而且木质化过程中,木质素的沉积也存在一定的规律性。利用红外光谱研究发现,木质素优先于纤维轴的取向沉积,沿纤维素基原纤维轴向排
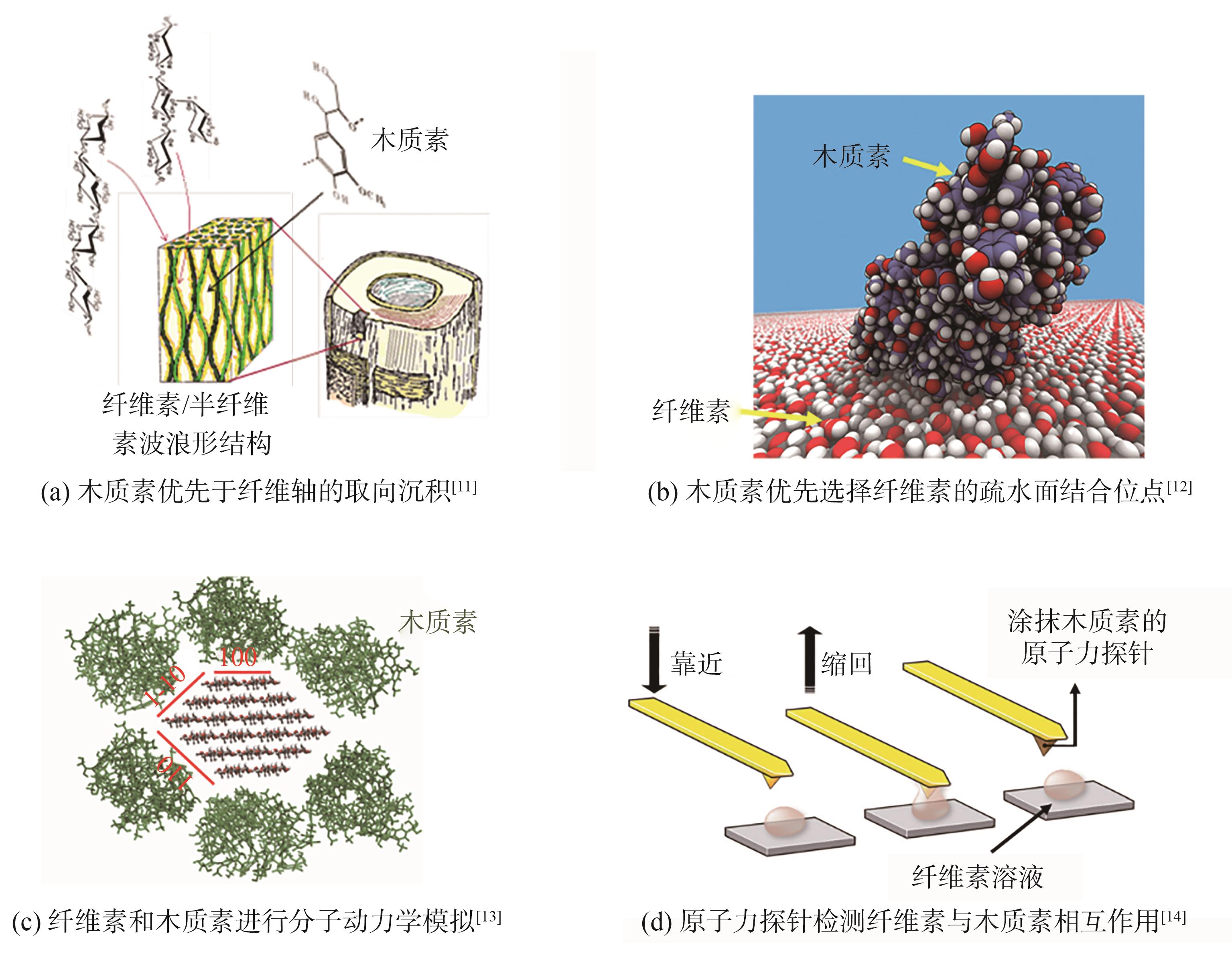
图3 纤维素与木质素的相互作用关系
Fig. 3 Interaction between cellulose and lignin
学者们采用分子模拟方法进一步研究了纤维素与木质素的相互作用关系(见
综上,木质素与纤维素的相互作用研究鲜见报道,有待进一步挖掘,特别需要关注二者之间的非共价键,以便于开发更加高效清洁的组分分离技术,提高生物质转化效率。
木质素与半纤维素间存在复杂的相互作用,包括共价键和非共价键,其中木质素与半纤维素通过共价键连接形成木质素-碳水化合物复合体(LCC),兼具木质素的机械强度、疏水性及多糖的生物相容性、抗致病性和亲水性等特征,是导致木质纤维生物质具有顽强的抗降解性的主要原
木质素是芳香族高分子化合物,而半纤维素是由非结晶复合聚糖组成的多分枝低分子聚合物,二者在细胞壁分化过程中通过生物合成、组分沉积的过程交联在一起,以不同方式及不同程度的共价键结合,形成LCC。学者们采用湿化学法对LCC进行分离及结构表征,认为木质素与半纤维素之间主要通过苄基醚键(BE)、苯基糖苷键(PG)、苯基酯键(Est)、γ-酯键(GE)及缩醛键(Acetal)等5种化学键连
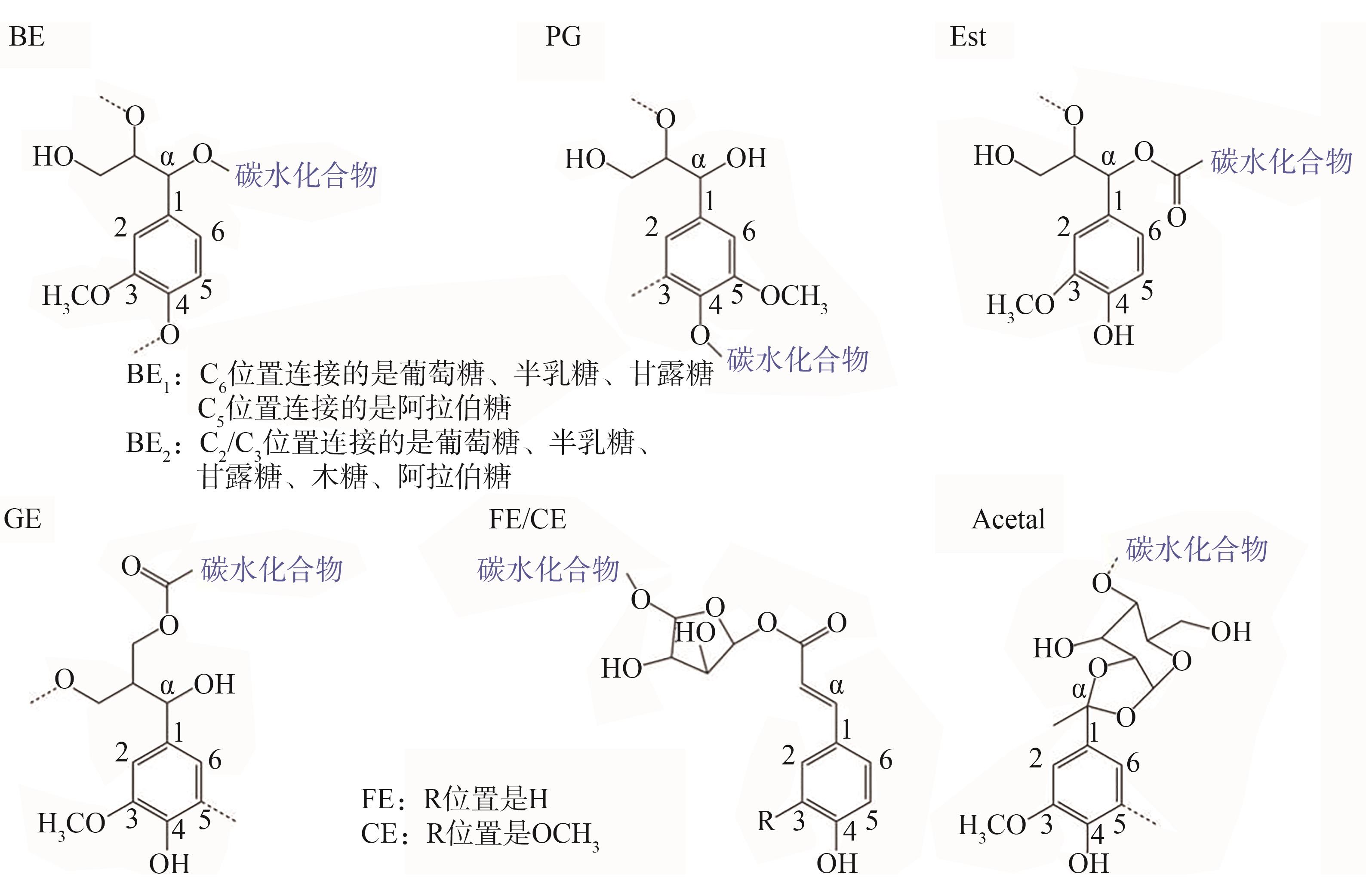
图4 木质素-碳水化合物之间的化学键类
Fig. 4 Chemical bond types of lignin-carbohydrat
不同植物纤维原料以及采用不同分离方法得到的LCC,其连接键不同。目前普遍认为,针叶木材中LCC的连接键主要是苄基醚键(落叶松可能存在苄基酯
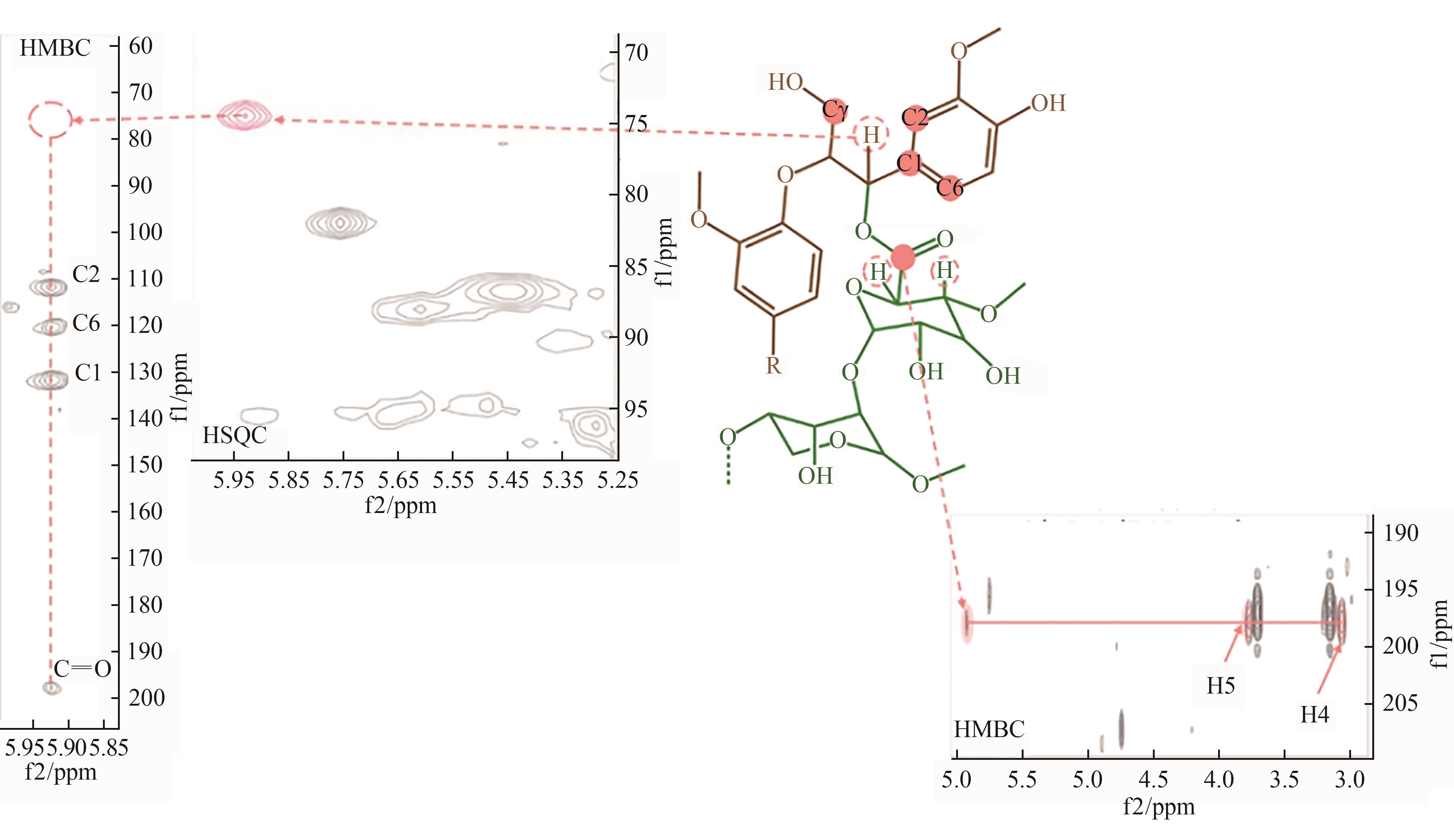
图5 木质素和阿拉伯葡萄糖醛酸聚木糖之间存在苄基酯
Fig. 5 Benzyl ester bonds between lignin and arabinaldehyde xyla

图6 不同方法分离的芦竹LCC HSQC谱
Fig. 6 HSQC spectra of LCC Arundo donax Linn isolated by different method
LCC中半纤维素与木质素的结合比例也不尽相同,据报道,针叶木材中几乎全部木质素与半纤维素结合,而阔叶木材中47%~66%的木质素与半纤维素结
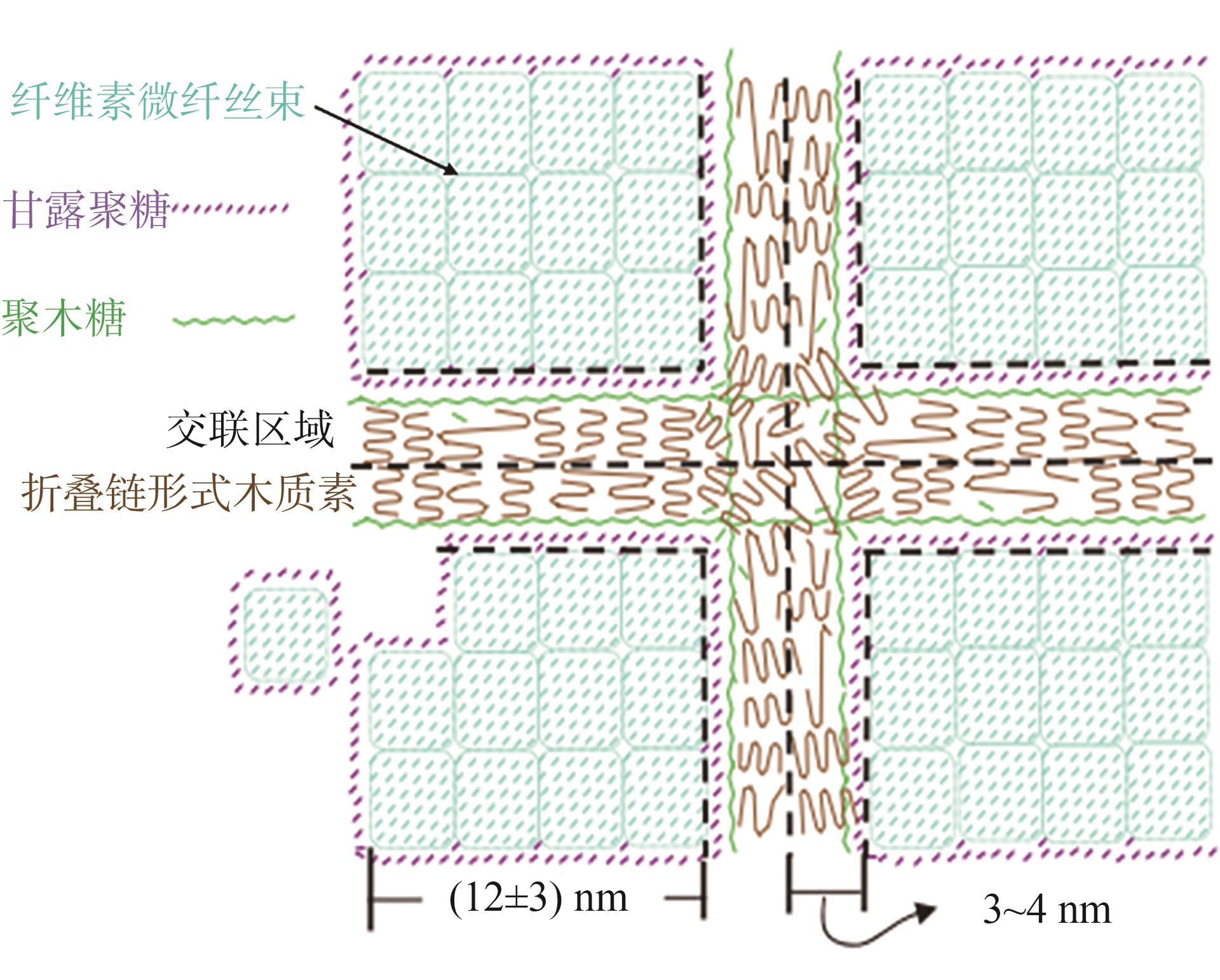
图7 木质素与半纤维素可能的相互作用关系示意
Fig. 7 Schematic diagram of possible interaction between lignin and hemicellulos
现有的LCC分离方法主要采用物理化学法,会破坏木质素和半纤维素原本的化学结构,导致其谱图信号改变或者丢失,为此,针对木质素与半纤维素之间相互作用的无损检测仍然是一个挑
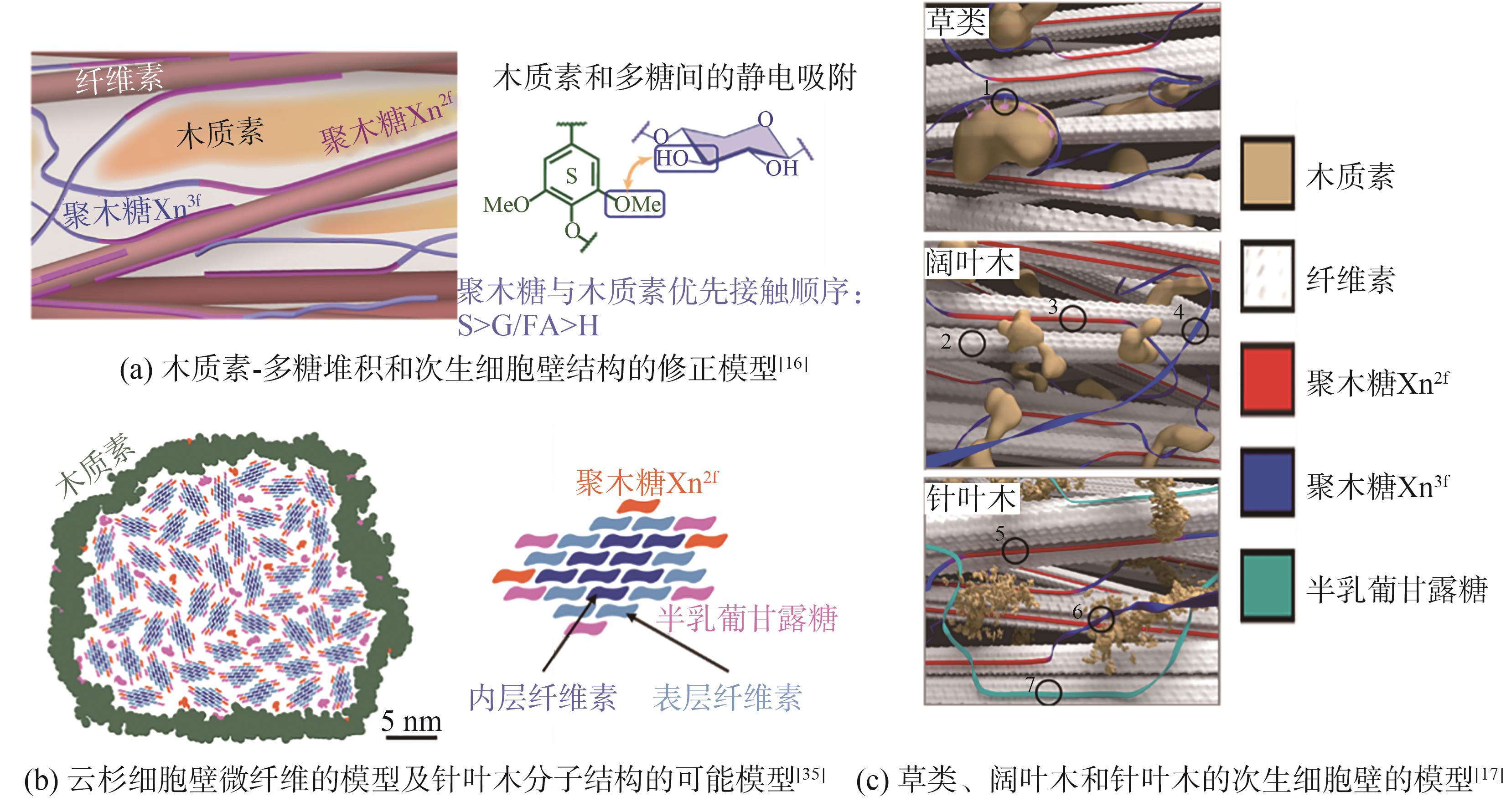
图8 植物细胞壁内的分子结构可能的模型
Fig. 8 Possible models of the molecular architecture of plant cell wall
木质素在禾本科纤维原料的细胞壁中与多糖存在广泛的非共价键,但在其他原料中是否存在相同的作用关系尚不明确。Terrett等
木质纤维生物质是加工生物基燃料、化学品和材料的重要可再生资源,但是由于其细胞壁具有抗降解性,导致生物基制品的生产过程存在高耗能、高排放及低效率等问题。了解木质纤维生物质的细胞壁复杂结构有利于通过基因工程定制植物或者优化加工技术以降低其抗降解性,对实现生物质高值化利用具有重要意义。近年来,人们在纤维素、半纤维素及木质素间相互作用方面的研究取得了良好进展,但是仍有一些科学问题待进一步解决。
(1)半纤维素结构复杂,特别是草类的聚木糖主链上存在支链和各类取代基,尽管已有大量研究表明取代基会影响聚木糖与细胞壁其他成分相互作用的方式,但是聚木糖上取代基的作用尚不清楚。
(2)由于表征手段限制,无法精确探测二螺旋结构聚木糖与纤维素表面结合的比例;木质素与纤维素的结晶区还是无定形区、与亲水面还是疏水面的结合等研究内容少有涉及。
(3)木质素与半纤维素间存在共价键和氢键,使得木质纤维生物质对酶促及微生物水解等作用具有很强的抗逆性,其中木质素与半纤维素相互作用关系(特别是化学键键合机制)尚未阐释清楚,值得以原位分析手段进一步深入探索。
总之,为了有效开发利用木质纤维生物质,需要利用先进的手段了解关于细胞壁内分子间相互关系的新知识,比如通过固体核磁共振技术、拉曼光谱、电子显微技术联用等方法,以期从分子层面上开启细胞壁复杂结构研究的新视角,为实现更高效的生物质精炼技术提供一定的理论支撑。
参考文献
PURUSHOTHAM P, HO R, ZIMMER J. Architecture of a catalytically active homotrimeric plant cellulose synthase complex[J]. Science, 2020, 369(6507): 1089-1094. [百度学术]
PHYO P, WANG T, YANG Y, et al. Direct determination of hydroxymethyl conformations of plant cell wall cellulose using
WANG T, YANG H, KUBICKI J D, et al. Cellulose structural polymorphism in plant primary cell walls investigated by high-field 2D solid-state NMR spectroscopy and density functional theory calculations[J]. Biomacromolecules, 2016, 17(6): 2210-2222. [百度学术]
BUSSE-WICHER M, LI A, SILVEIRA R L, et al. Evolution of xylan substitution patterns in gymnosperms and angiosperms: Implications for xylan interaction with cellulose[J]. Plant Physiology, 2016, 171(4): 2418-2431. [百度学术]
GRANTHAM N J, WURMAN-RODRICH J, TERRETT O M, et al. An even pattern of xylan substitution is critical for interaction with cellulose in plant cell walls[J]. Nature Plants, 2017, 3(11): 859-865. [百度学术]
SIMMONS T J, MORTIMER J C, BERNARDINELLI O D, et al. Folding of xylan onto cellulose fibrils in plant cell walls revealed by solid-state NMR[J]. Nature Communications, DOI: 10.1038/ncomms13902. [百度学术]
ZUK M. Shallow ponds prompt fitness-favorable species interbreeding[J]. Science, 2020, 367(6484): 1304-1305. [百度学术]
RALPH J, BRUNOW G, HARRIS P J, et al. Lignification: Are lignins biosynthesized via simple combinatorial chemistry or via proteinaceous control and template replication? [M]//Lattanzio V, Daayf F. Recent Advances in Polyphenol Research. Oxord: Wiley, 2009: 36-66. [百度学术]
Gao Y, Lipton A S, Wittmer Y, et al. A grass-specific cellulose-xylan interaction dominates in sorghum secondary cell walls[J]. Nature Communications, DOI: 10.1038/s41467-020-19837-z. [百度学术]
ÅKERHOLM M, SALMÉN L. The oriented structure of lignin and its viscoelastic properties studied by static and dynamic FT-IR spectroscopy[J]. Holzforschung, 2003, 57(5): 459-465. [百度学术]
SALMÉN L, OLSSON A, STEVANIC J S, et al. Structural organisation of the wood polymers in the wood fibre structure[J]. Bioresources, 2012, 1(7): 521-532. [百度学术]
BESOMBES S, MAZEAU K. The cellulose/lignin assembly assessed by molecular modeling. Part 2: Seeking for evidence of organization of lignin molecules at the interface with cellulose[J]. Plant Physiology and Biochemistry, 2005, 43(3): 277-286. [百度学术]
VERMAAS J V, CROWLEY M F, BECKHAM G T. A quantitative molecular atlas for interactions between lignin and cellulose[J]. ACS Sustainable Chemistry & Engineering, 2019, 7(24): 19570-19583. [百度学术]
ZHANG B, YANG J, LIU Y, et al. Effect of temperature on the interactions between cellulose and lignin via molecular dynamics simulations[J]. Cellulose, 2022, 29(12): 6565-6578. [百度学术]
NASCIMENTO D M, COLOMBARI F M, FOCASSIO B, et al. How lignin sticks to cellulose-insights from atomic force microscopy enhanced by machine-learning analysis and molecular dynamics simulations[J]. Nanoscale, 2022, 14(47): 11757-17561. [百度学术]
KANG X, KIRUI A, DICKWELLA WIDANAGE M C, et al. Lignin-polysaccharide interactions in plant secondary cell walls revealed by solid-state NMR[J]. Nature Communications, DOI:10.1038/s41467-018-08252-0. [百度学术]
KIRUI A, ZHAO W, DELIGEY F, et al. Carbohydrate-aromatic interface and molecular architecture of lignocellulose[J]. Nature Communications, DOI: 10.1038/s41467-022-28165-3. [百度学术]
ZHAO Y, SHAKEEL U, SAIF UR REHMAN M, et al. Lignin-carbohydrate complexes (LCCs) and its role in biorefinery[J]. Journal of Cleaner Production, DOI: 10.1016/j.jclepro.2020.120076. [百度学术]
SAPOUNA I, LAWOKO M. Deciphering lignin heterogeneity in ball milled softwood: Unravelling the synergy between the supramolecular cell wall structure and molecular events[J]. Green Chemistry, 2021, 23(9): 3348-3364. [百度学术]
JEONG S, LEE E, BAN S, et al. Structural characterization of the lignin-carbohydrate complex in biomass pretreated with Fenton oxidation and hydrothermal treatment and consequences on enzymatic hydrolysis efficiency[J]. Carbohydrate Polymers, DOI: 10.1016/j.carbpol.2021.118375. [百度学术]
SU C, GAN T, LIU Z, et al. Enhancement of the antioxidant abilities of lignin and lignin-carbohydrate complex from wheat straw by moderate depolymerization via LiCl/DMSO solvent catalysis[J]. International Journal of Biological Macromolecules, 2021, 184: 369-379. [百度学术]
WANG X, HAN J, PANG S, et al. Structural enrichment and identification of lignin-carbohydrate complex in alkaline stabilized system[J]. Carbohydrate Polymers, DOI: 10.1016/j.carbpol.2022.119873. [百度学术]
ZHAO J, HAN J, CHEN Z, et al. Structural evolution of lignin after selective oxidation of lignin-carbohydrate complex by chlorine dioxide[J]. International Journal of Biological Macromolecules, 2022, 223: 273-280. [百度学术]
GIUMMARELLA N, LAWOKO M. Structural insights on recalcitrance during hydrothermal hemicellulose extraction from wood[J]. ACS Sustainable Chemistry & Engineering, 2017, 5(6): 5156-5165. [百度学术]
YOU T, ZHANG L, ZHOU S, et al. Structural elucidation of lignin-carbohydrate complex (LCC) preparations and lignin from Arundo donax Linn[J]. Industrial Crops and Products, 2015, 71: 65-74. [百度学术]
ZHAO B, CHEN B, YANG S, et al. Structural variation of lignin and lignin-carbohydrate complex in Eucalyptus grandis × E. urophylla during its growth process[J]. ACS Sustainable Chemistry & Engineering, 2017, 5(1): 1113-1122. [百度学术]
路德胜, 张凯, 陈旭东, 等.
LU D S, ZHANG K, CHEN X D, et al. Study on the Structure of Residual Lignin in Unbleached Kraft Pulp by
BALAKSHIN M, CAPANEMA E, GRACZ H, et al. Quantification of lignin-carbohydrate linkages with high-resolution NMR spectroscopy[J]. Planta, 2011, 233(6): 1097-1110. [百度学术]
LAWOKO M, HENRIKSSON G, GELLERSTEDT G. Structural differences between the lignin-carbohydrate complexes present in wood and in chemical pulps[J]. Biomacromolecules, 2005, 6(6): 3467-3473. [百度学术]
HENRIKSSON G, LAWOKO M, MARTIN M E E, et al. Lignin-carbohydrate network in wood and pulps: A determinant for reactivity[J]. Holzforschung, 2007, 61(6): 668-674. [百度学术]
TERASHIMA N, KITANO K, KOJIMA M, et al. Nanostructural assembly of cellulose, hemicellulose, and lignin in the middle layer of secondary wall of ginkgo tracheid[J]. Journal of Wood Science, 2009, 55(6): 409-416. [百度学术]
李海潮, 罗宇欣, 苗国华, 等. 全生物基三元DES预处理促进芦竹酶解机理研究[J].中国造纸学报,2022,37(4):31-38. [百度学术]
LI H C, LUO Y X, MIAO G H, et al. Study on Mechanism of lmproving Enzymatic Hydrolysis of Arundo donax L. Pretreated with Bio-based Ternary DES[J]. Transactions of China Pulp and Paper, 2022, 37(4): 31-38. [百度学术]
LAWOKO M, VANHEININGEN A R P. Fractionation and characterization of completely dissolved ball milled hardwood[J]. Journal of Wood Chemistry and Technology, 2011, 31(3): 183-203. [百度学术]
CHENG K, SOREK H, ZIMMERMANN H, et al. Solution-state 2D NMR spectroscopy of plant cell walls enabled by a dimethylsulfoxide-d6/1-ethyl-3-methylimidazolium acetate solvent[J]. Analytical Chemistry, 2013, 85(6): 3213-3221. [百度学术]
TERRETT O M, LYCZAKOWSKI J J, YU L, et al. Molecular architecture of softwood revealed by solid-state NMR[J]. Nature Communications, DOI: 10.1038/s41467-019-12979-9. [百度学术]